|
|
||||
 |
 |
|
 |
 |
|
|
||||
|
|
||||
|
|
|
|||
|
|
Atlas of Cyanobacteria https://atlasofcyanobacteria.myrevbase.com/v
|
|
||
Cyano-
= pigment color in the blue-green part of the visible spectrum due to the
presence of light-capturing molecules including chlorophyll a (green) as well
as water-soluble 'phycobilins' phycoerithrin (red), phycocyanin and
allophycocyanin (blue), and other accessory yellow pigments, predominantly
β-carotene and zeaxanthin (Hirschberg and Chamovitz 2004). Color varies
from nearly black to colorless.
Old
Names: Myxophyceae, Schizophyceae (schizo- =
split), Schizomycetes and Myxobacteria (myxo- = slime) are retired names
(Geitler 1932), as is Cyanophyceae used by Geitler himself (1932, 1979). The name ‘blue-green algae’ appeared as
recently as 1971 by Stanier et al. (1971). Other retirees include 'Cyanophycota' (Friedmann 1982) and
'Blue-greens' (Fay 1983). 'Cyanophyta' and 'blue-green algae' appeared again
recently (van den Hoek et al. 2005; Whitton 2011) as equivalent to
Cyanobacteria while explicitly describing their bacterial structure.
Bacteria: Because most cyanobacteria are photosynthetic and often share the same habitat as eucaryotic photosynthetic protists ("algae"), they were previously classed as cyanophyceae, where “‑phyceae” in botanical nomenclature signifies eukaryotic “algae”. Roger Stanier (1916 - 1982) stated clearly (ca. 1970) that these photosynthetic bacteria should be classified according to the rules of procaryote nomenclature, published in later papers ( Stanier et al. 1971, Stanier 1977), hence their currently accepted name. In bacterial classification only living cultures are recognized as “holotypes” capable of being distinguished, in contrast to the “type specimen” (dead; not cultures) required in the Botanical Code. Thus revisions are occurring both at the generic and species level (Ripka et al. 1978). Polymorphism: Without molecular information the identity of species and in some case genera is doubtful. Ecads, or morphotypes modified by the microhabitat, modify sizes, shapes and both intensity and color of pigmentation. The viscosity of extracellular glycans (polysaccharides with various structure) changes the shape and orientation of coccoid cells as well as filaments. “In no class have there been so many records of polymorphism as in the Cyanophyceae” (West and Fritsch 1927). Photopigments vary in
dominance and ratio, but generally include Chl a, phycocyanin, allophycocyanin, phycoerithrin and carotenoids. As a result
the apparent cytoplasmic color also varies from red to green to cyan to blue, as well as brown and essentially black.
Such variation may be influenced by the light available in any particular
microhabitat, such as in the microstratified layers of a thermocline --
chromatic adaptation.
Photosynthetic pigment absorbance spectra of cyanobacteria (euryaerobic - from anaerobes to aerobes), purple bacteria (microaerobes), and green bacteria (strict anaerobes). © 2007 Sinaur Associates posted as slide 19 online. Chromatic adaptation is the ability of cyanobacteria, especially of similar strains, to vary their pigment ratio, especially the accessory phycobilins (allophycocyanin, phycocyanin, and phycoerithrin. A clear example is the change in ratio of phycocyanin to phycoerithrin in Synechoccus in chemostats, both in uniculture and mixed culture, when confronted with either green or red light. the red strain (rich in phycoerithrin) outcompeted the green strain in green light, and vice versa (Stomp et al. 2004). Effectively this reduces inter-strain competition, and increases diversity of strains by complementarity – absorbing different light spectra when confronted with light shading by one of the strains. Other more remotely related strains such as Tolypothrix can also compensate for changes in available light, as found in mixed cultures with the red or green strain of Synechococcus (Ibid.). Another
example: As a graduate student at the University of Minnesota I kept a 500
ml capped bottle of metalimnetic Oscillatoria
agardhii (now Planktothrix
agardhii) in the laboratory at room temperature, with only the overhead fluorescent
fixtures as a light source, and without any disturbance such as nutrient
addition. Within a few weeks I was astounded to see that it had turned red,
and it would have been called O.
rubescens. To this day I wonder whether there were a few scarce
red filaments in the lakewater, or whether the green trichomes turned red. I
searched for some half-green half-red trichomes but found none...and never
saw red trichomes in the source -- Deming Lake, Itasca State Park, Minnesota
(47o 10' 13.66" N, 95o 10' 07.18" W).
Confirmation of this 'duality' is clear: Although allozymes (a.k.a. alloenzymes -- variant forms of an enzyme that are coded by different alleles at the same locus are identical, absorbance spectra of Oscillatoria rubescens (1) and O. agardhii are different (Komárek 2000, Figure 8):
No flagella are
present, but some "gliding” or “bending (flexing)” motility occurs in
several filamentous genera lacking dense outer sheaths associated with
individual trichomes. Good examples include Oscillatoria (some having been renamed
"Planktothrix"), Arthrospira,
and Microcoleus (with a common sheath containing multiple trichomes not firmly fixed in the
sheath). The motility can take the form of reversible translocation along the
axis of the trichome, generally with clockwise or counterclockwise rotation.
In other cases the motility is pendulum-like bending or flexing of the axis
of the trichome. In Lyngbya,
hormogones (short fragments containing a few cells) can be seen travelling
along the inside of an empty sheath, eventually escaping to form their own
filament.
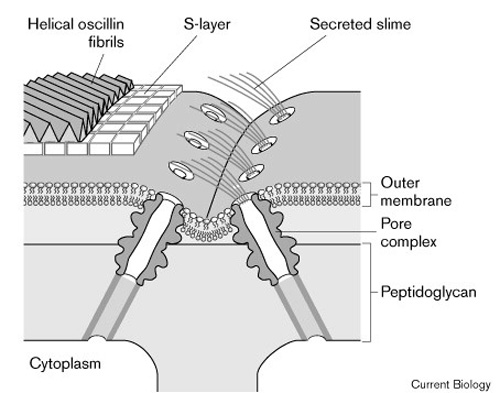
Motility mechanism: Direct observation of mucilage secretion
(Hoiczyk and Baumeister 1998) during gliding motility of unbranched
trichomes, including mucilage-secreting pores and mucilage trails, along with
electron micrographs of the pores at trichome cell junctions, enabled a model
in which helical proteins in the outer cyanobacterial wall control secretion
of mucilage to propel the trichome reversibly, with a reversing rotation and
translocation. How all the cells in the trichome are communicating to act
synergically (to propel reversibly) is not explained. Also, trichome bending
may require a different model. How external stimuli (light, temperature,
solutes) are sensed by the motility system is another series of questions.
Origin and Age: The oldest known fossils are from the Archaean Era, ~3.5 Ga (billion years), in stromatolites found in Western Australia and South Africa. Cyanobacteria along with other bacteria and archaea likely were the first life forms to inhabit Earth. Their ultimate origin may well be extraterrestrial given their extraordinary ability (at least in spore form) to resist extremes of low temperature, low moisture, and capacity (at least some genera) for both anaerobic and aerobic metabolism. Thus as a group they may be older than Earth itself, currently estimated to have accreted 4.5 Ga. Given ca. 1 billion years of cooling to a temperature tolerable to terrestrial life, and the relative complexity both morphologically and physiologically, it is not unreasonable to expect extraterrestrial procaryotes to land on Earth, and perhaps distant hospitable planets outside the Solar System, and develop successful communities. Importance in Carbon Fixation: Cyanobacteria currently and collectively account for >30% of global photosynthesis primarily as oceanic picoplankton (Rae et al.), slowing (mitigating) both the rise in atmospheric CO2 and rate of ocean acidification. Importance
in Oxygen Fixation: By way of endocytosis, a unicellular cyanobacterium captured and retained by a colorless eukaryote evolved into a chloroplast, hence was the origin of all protistan and plant chloroplasts (Margulis 1967).
Importance in Nitrogen Fixation: Along with several other photosynthetic and heterotrophic bacteria, the cyanobacteria can reduce elemental nitrogen (unavailable biologically except in this way) to ammonia - 'nitrogen fixation', the source of nitrogen for amino acids and proteins. Grula (2005) suggests that N-fixation was essential to early cyanobacteria, and evolved either before or at the same time as synthesis of chlorophyll. N-fixation is an anoxic reaction but can occur in oxic environments simultaneous with oxic photosynthesis within separate intrabacterial compartments. In heterocystous cyanobacteria such as Anabaena spp., the separate compartment is the heterocyst. An unusual case of N-fixation occurs in the unicellular picocyanobacterium Candidatus (uncultured to date) Atelocyanobacterium thalassa (a.k.a. UCYN-A) loosely connected as an epiphyte on the prymnesiophyte Braarudosphaera bigelowii, a coastal marine coccolith found at high latitudes. The relationship is thought to be an example of obligate symbiosis. A. thalassa lacks PS II (no O2 production and apparently no glucose production) and some other metabolic pathways, but does fix nitrogen. B. bigelowii is eukaryotic and photosynthetic, and produces pentagonal calcareous scales. Stable isotope tracer experiments demonstrate that the cyanobacterium provides fixed nitrogen to the eucaryote, which in turn provides glucose to the bacterium. (Thompson et al. 2012.) Importance in Methane Production: Classically methane production has been relegated to the archaea, specifically the methanogens, restricted to anoxic environments such as sediments and overlying anoxic water zones. Accumulating evidence of methane in oxic water layers has given rise to the 'methane anomoly' that questions where and how the methane is produced. Bižić et al. (2020) have unequivocal evidence that several unicellular and filamentous cyanobacteria produce methane by way of an undefined light-driven methanism requiring a minimum of 20 mmol quanta m−2 s−1. Methane was produced under each of light, dark, oxic and anoxic conditions by freshwater, marine and terrestrial cyanobacteria, including some major bloom formers such as Microcystis aeruginosa (freshwater lake plankton) and Trichodesmium erythraeum (marine plankton). Additional evidence that excluded activirty of archaea included use of several axenic cyanobacterial cultures, lack of necessary genes and/or lack of their expression, and stable isotope utilization (13C-labeled sodium bicarbonate). The impact of surficial aerobic methanogenesis is clearly its contribution to the total atmospheric load of greenhouse gasses and warming of the biosphere. Methane has far more global warming potential (GWP) than carbon dioxide. CH4 has a relatively short lifetime in the atmosphere. The 100-year GWP of 28–36 and the 20-year GWP of 84–87 indicate the more powerful impact of energy absorption relative to CO2 with a GWP value of 1. This is the case even though CO2 has a mass emision of 81% and CH4 is only 10% of greenhouse gasses. (For comparison, Nitrous oxide (N2O) with only 7% mass emission of greenhouse gasses is the worst offender with a GWP 265–298 for a 100-year timescale.) See: https://www.epa.gov/ghgemissions/overview-greenhouse-gases
The
spheroid bodies are now known to have well conserved genes for nitrogen
fixation, and lack one or more genes for production of chlorophyll. In a comparison of spheroid bodies in the
diatoms Rhopalodia gibberula and Epithemia turgida, those in the R. gibberula retained most genes for chlorophyll production, while those in E. turgida had
none. Similarly genes for production
of vitamin B12 were intact in the R. gibberula bodies but almost entirely
missing in E. turbida bodies (Nakayama and Inagaki 2017).
Cyanobacterial
endosymbionts have been found in several genera of non-photosynthetic dinophyceae,
dictyophyceae (silicoflagellates) and tintinnids (a group of marine ciliates)
that are genetically close to Synechococcus and Prochlorococcus (Foster et al.
2006).
Toxins:
|
||||
Secondary metabolites (not required for growth) include nitrogen (N)-rich substances that are most likely a reserve for N but at the same time are toxic to humans, other mammals and likely other organisms. Physiologically they are known to degrade either mammalian livers (hepatotoxins) or nerve networks (neurotoxins). Nearly all lakes have toxic cyanobacteria, even the most oligotrophic in New Hampshire USA also contain toxins such as microcystin, a liver toxin (Haney 19XX) including the genera Microcystis and Oscillatoria (Planktothrix). Evidence that the toxins have diminished production in extreme lakes with > 2000 µg total nitrogen (TN) is in Midwestern lakes in the USA (Graham et al. 2004).
From Graham et al. 2004, Figure 2a. The lack of toxin production where N is in such excess reinforces the idea that they are primarily a N-storage mechanism.
|
||||
Blooms: |
||||
A large concentration of biologically available phosphorus and limited concentrations of nitrate-N tend to favor high growth rates of N-fixing cyanobacteria with or without heterocysts, relative to other PS plankton. Buoyant cyanobacteria accumulate at the water surface forming blooms. Generally a period of rapid growth and population increase occurs well below the surface and beneath the impact of injurious UV radiation, and bloom formation can mark the end of the growth of a clone. A series of several blooms may follow each other as subsequent clonal populations develop sequentially, ending their pulse by floating to the surface. Wind can mix the blooms downward dispersing the cyanobacteria, or at lower velocity can concentrate the potentially toxic populations along the lee shores providing a danger to vertebrates. Chemical control of cyanobacterial blooms in New Hampshire USA, at least until ~1980, included CuSO4 solution added to lake surfaces either by spraying or by dragging crystals in hemp sacks suspended from a power boat in transects across lakes. Massive fish kills often followed copper treatments, largely from anoxia and toxin release as the cyanobacteria lysed. Recognition that copper toxicity was not limited to the cyanobacteria, the temporary nature of the bloom control and high costs have made the application unpopular in the state. An alternative chemical control is application of alum, a co-precipitate of Al2(SO4)3 and Na2Al2O4 (or related salts) that adsorbs phosphate from the water column and carries it to the sediments. Expense, limited duration and potential aluminum toxicity limit its use. Biological control may have at least a minimal impact on cyanobacterial growth prior to blooms. Simocephalus vetulus, a large 2 – 4 mm long caldoceran, can grow on diets of some strains of Microcystis (but not all) and Oscillatoria (“Limnothrix”) sp. as well as on the green Scenedesmus acutus (Fernández et al. 2014). Some ciliates and amoebae also consume cyanobacteria but are unlikely to prevent blooms. Cyanobacteria in Aerosols
Bacteria
and eukaryotic protists as well as non-living
particles in soils and water can become airborne as ‘aerosols’. Energy for launching is lower from dry
soils than from wet soils or water, where bubble bursts and evaporation
occur.
I
propose that the epilimnion in relatively calm lakewater is effectively a dry surface onto which dry particles in the same size range
as unicells including some cyanobacteria can settle
without penetrating the water surface. Small air updrafts should lift them back into the atmosphere about as
easily as from dry soil surfaces. If
an insect as large as the Water Strider (Gerris spp.) can walk and even jump from the epineuston ‘skin’ without getting wet, certainly bacterial-sized particles can also.
One
of several factors related to aerosol liberation from water includes
viscosity, itself a function of temperature and even bacterial concentration. Superfluid conditions can be created with a concentration of ~ 5x109 flagellated bacteria per ml with rheology rotating
probe in a container) measurements (López et al.
2015).
Aerosols
are known to migrate great distances around the globe, including 55 types of viable
cyanobacteria and a total of 187 viable airborne algae and protrozoa collected over Germany, Netherlands, Taiwan and
the U.S.A. (Schlichting 2012). Several cyanobacteria including Nostoc,
Oscillatoria, Calothrix and Synechococcus have been found based on SSU rRNA at altitudes of
up to 10 km in the upper Troposphere (DeLeon-Rodriguez
et al. 2013). Cyanobacteria, protists and other small particles are also carried by waterfowl as shown by samples
taken from ducks suspended in diapers on a clothesline for at least two weeks
(Schlichting 1958). Unique sampling methods included a radio-controlled electric boat (Schlichting and Hudson 1967) and piloting an open cockpit
biplane holding agar plates in the air while circling the football stadium at
Michigan State University (personal communication).
If
cyanobacterial toxins such as microcystins, saxitoxins and others are a direct cause of mammalian pathology including hepatoxins and neurotoxins, there are consequences of
inhaling aerosols containing them. Schlichting (2012) estimated the
average human inhales ~ 7 liters of atmosphere per day including ~ 2880
cyanobacteria). Such aerosols are found
in the outside environment as well as inside office buildings (Chu et al.
2013).
Predators on cyanobacteria The
chrysophycean mixotrophic flagellate Ochromonas can feed on all Microcystis strains
tested as well as on Pseudanabaena,
and have a ‘strongly reduce’ Microcystis biomass and toxins”, thus
potentially reducing severity of blooms, or at least reduce the frequency of
small colonies (≤ 810 µm3)
as demonstrated in culture. Ochromonas and Microcystis co-occurred in 94% of 460 Norwegian lakes. Apparently Ochromonas digests ingested cyanobacterial cells or even ingested
dissolved microcystin (van Donk et al. 2009). There is evidence that Ochromonas can be amoeboid, producing pseudopods to capture particles (Boenigk and
Arndt 2000).
Natural populations of filamentous cyanobacteria such as Oscillatoria (a.k.a. Planktothrix) can also be decimated by ‘gulping’ ciliates such as Pseudomicrothorax dubius (Hausmann 2002). Logically the best control is preventative: Reduce phosphorus loading from the watershed.
|
||||
|
References: |
||||
Boenigk,
J. and H. Arndt 2000. Particle handling during interception
feeding by four species of heterotrophic nanoflagellates. Journal of Eukaryotic
Microbiology, 47,
350–358.
Burke,
D. H., Hearst, J. E. & Sidow, A. (1993). Early evolution of
photosynthesis: clues from nitrogenase and chlorophyll iron proteins. Proc. Natl. Acad. Sci. USA 90, 7134-7138.
Bižić, M., T. Klintzsch, D. Ionescu1, M. Y. Hindiyeh, M. Günthel, A. M. Muro-Pastor, Chu,
W-L., S-Y Tneh and S. Ambu 2013. A survey of airborne algae and
cyanobacteria within the indoor environment of an office building in Kuala
Lumpur, Malaysia.
DeLeon-Rodriguez,
N., T.L. Lathem, L.M. Rodrigues-R,
et al. 2013. Microbiome of the
upper troposphere: Species composition
and prevalence, effect of tropical storms, and atmospheric implications. Proceedings of the National Academy of
Sciences of the United States of America 110(7):2575-2580.
Donis, D., S. Flury, A. Stöckli, J. E. Spangenberg, D. Vachon, D. F. McGinnis. 2017 Full-scale Fay,
P. 1983. The Blue-greens. Edward Arnold, London.
Fernández, R., S. Nandini, S.S.S. Sarma and M.E. Castellanos-Paez 2014. Effects of cyanobacteria, fish kairomones, and the presence of ostracods on the demography of Simocephalus vetulus (Cladocera). Invertebrate Biology 133(4):371-380. Foster,
R., J.L. Collier & E.J. Carpenter 2006. Reverse transcription PCR
amplification of cyanobacterial symbiont 16S rRNA sequences from single
non-photosynthetic eukaryotic marine planktonic host cells. Journal of Phycology 42:243-250.
Friedman,
I. 1982. Cyanophycota. In:
Synopsis and Classification of Living Organisms, Vol. I, Ed. S.P. Parker, pp.
45-52. McGraw-Hill, NY.
Geitler, L. 1932. Cyanophyceae. Johnson Reprint Corporation, NY, London. [Cyanophyceae von Europe, a part of Rabenhorst’s Kryptogamen-Flora von Deutschland, Österreich und der Schweiz.] Geitler, L. 1977. Zur Entwicklungsgeschichte der Epithemiaceen Epithemia, Rhopalodia und Denticula (Diatomophyceae) und ihre vermutlich symbintischen Sphäroidkörper. Plant Syst. Evol. 128:295–275. Geitler, L. 1979. Einige kritische Bemerkungen zur neuen zusammenfassenden Darstellung der Morphologie und Systematik der Cyanophyceen. Plant Syst. Evol. 132:153-60. Graham, J.L., J.R. Jones, S.B. Jones, J.A. Downing and T.E. Clevenger 2004. Environmental factors influencing microcystin distribution and concentration in the Midwestern United States. Water Research 38:4395-4404. (online) Grula, J.W. 2005.
Evolution of photosynthesis and biospheric oxygenation contingent upon nigroten fixation? Internat. J. of Astrobiology 4:251-257. (online)
Hausmann, K. 2002. Food acquisition, food ingestion and food digestion by protists. Japanese Journal of Protozoology 35(2): 85 – 95. Hirschberg,
J., and D. Chamovitz 2004. Carotenoids in
cyanobacteria. Advances in Photosynthesis and Respiration 1:559-579.
Hoiczyk, E., and W. Baumeister 1998. The junctional pore complex, a prokaryotic secretion organelle, is the molecula motor underlying gliding motility in cyanobacteria. Current Biology
8:1161-1168
Lockhart, P.J., A.W.D. Larkum, M.A. Steel, P.J. Waddell, and D. Penny 1996. Evolution of chlorophyll and bacteriochlorophyll: The problem of invariant sites in sequence analysis. Proc. Natl. Acad. Sci. USA 93:1930-1934. López, H.M., J. Gachelin, C. Douarche, H. Auradou and E. Clément 2015. Turning bacteria suspensions into superfluids. Physical Review Letters 115:1-5. Margulis, L. 1967. On the origin of mitosing cells. Journal of Theoretical Biology 14(3):225-274. Matthews, R.A. 2016. Volume 1, Cyanobacteria. Books and Monographs. Book 6. http://cedar.wwu.edu/cedarbooks/6 Nakayama, T. & Y. Inagaki 2017. Genomic divergence within non-photosynthetic cyanobacterial endosymbionts in rhopalodiacean diatom. Scientific Reports 7, Article number 13075. Prechtl, J., C. Kneip,
P. Lockhart, K. Wenderoth & U-G. Maier 2004. Intracellular spheroid bodies of Rhopalodia gibba have
nitrogen-fixing apparatus of cyanobacterial origin. Molecular Biology and Evolution 21(8):1477-1481.
Rae, B.D., B.M. Long, L.F. Whitehead, B. Förster, M.R. Badger, G.D. 2013. Cyanobacterial carboxysomes: Microcompartments that facilitate CO2 fixation. J. Mol. Microbiol. Biotechnol.23: 300 - 307. Rippka, R., J. Deruelles,
J.B. Waterbury, M. Herdman and R.Y. Stanier 1978. Generic
assignments, strain histories and properties of pure cultures of
cyanobacteria. Journal of General Microbiology 111:1 - 61.
Schlichting,
H. E., Jr., 1958. The Role of
Waterfowl in the Dispersal of Algae, Ph.D. Thesis, Mich. State Univ. 259 pp.,
Univ. Microfilm, Ann Arbor, Mich.
Schlichting,
H.E., Jr., 1961. Viable species of
algae and Protozoa in the atmosphere, Lloydia., 24: 2, 81-88.
Schlichting,
H.E., Jr., 1964, Meteorological conditions affecting the dispersal of airborne
algae and Protozoa. Lloydia, 27: 1, 64-78.
Schlichting,
H.E., Jr. and J.E. Hudson, Jr. J. E., 1967. "Radio-controlled model boat
samples air and plankton. Science 156:
3772, 238-239.
Schlichting,
H.E. Jr., 2012. The importance of airborne algae and protozoa.
Stanier, R.Y., R. Kunisawa, M. Mandel, and G. Cohen-Bazire 1971. Purification and properties of
unicellular blue-green algae (Order Chroococcales). Bacterial Rev. 35:171-205.
Stanier, R.Y. 1977. The position of the Cyanobacteria in the world of phototrophs. Carlsberg Res. Comm. 42:77-98. Stomp,
M., J. Huisman, F. de Jongh,
A.J. Veraart, D. Gerla,
M. Rijkeboer, B.W. Ibellings,
U.I.A. Wollenzien, and L.J. Stal. 2004. Adaptive divergence in pigment composition promotes phytoplankton
biodiversity.
Thompson, A., R.A. Foster, A. Krupke, B.J. Crter, N.Musat, D. Vaulot, M.M.M. Kuypers, and J.P. Zehr. 2012. Unicellular Cyanobacterium Symbiotic with a Single-Celled Eukaryotic Alga. Science 21 Vol. 337, Issue 6101: 1546-1550. van den Hoek, C., D.G. Mann, and H.M. Jahns 1995. Algae: an introduction to phycology.
Cambridge University Press (623 pp).
Whitton, B.A. 2011. Phylum Cyanobacteria (Cyanophyta). In: John, D.M., B.A. Whitton, and A.J. Brook (Eds.) The Freshwater Algal Flora of the British Isles. (878 pp)
UPDATE 9 Feb. 2021
|
||||
|
||||



 Modified from Komárek 2000, Figure 8.
Modified from Komárek 2000, Figure 8.